The structure of a pollen grain. The structure of a pollen grain Pollen grains in angiosperms are formed in
A flower is a conspicuous, often beautiful, important part of flowering plants. Flowers can be large or small, brightly colored and green, fragrant or odorless, solitary or collected together from many small flowers into one common inflorescence.
A flower is a modified shortened shoot used for seed propagation. The main or side shoot usually ends in a flower. Like any shoot, a flower develops from a bud.
Flower structure
A flower is the reproductive organ of angiosperms, consisting of a shortened stem (flower axis), on which the flower cover (perianth), stamens and pistils, consisting of one or more carpels, are located.
The axis of the flower is called receptacle. The receptacle, growing, takes different shape flat, concave, convex, hemispherical, cone-shaped, elongated, columnar. The receptacle below turns into a peduncle, connecting the flower with the stem or peduncle.
Flowers without a pedicel are called sessile. On the peduncle of many plants there are two or one small leaves - bracts.
Cover of a flower - perianth- can be divided into a calyx and corolla.
Cup forms the outer circle of the perianth, its leaves are usually relatively small sizes, Green colour. There are separate and fused calyx. Usually it serves the function of protecting the internal parts of the flower until the bud opens. In some cases, the calyx falls off when the flower opens; most often it remains during flowering.
The parts of the flower located around the stamens and pistil are called the perianth.
The inner leaflets are the petals that make up the corolla. The outer leaves - sepals - form a calyx. The perianth, consisting of a calyx and corolla, is called double. A perianth that is not divided into a corolla and a calyx, and all the leaflets of the flower are more or less the same - simple.

whisk — inner part perianth, differs from the calyx in its bright color and larger size. The color of the petals is due to the presence of chromoplasts. There are separate and fused corollas. The first consists of individual petals. In fused-petal corollas, a tube is distinguished and a limb located perpendicular to it, which has a certain number of teeth or corolla blades.
Flowers can be symmetrical or asymmetrical. There are flowers that do not have a perianth; they are called naked.
Symmetrical (actinomorphic)- if many axes of symmetry can be drawn through the rim.
Asymmetrical (zygomorphic)- if only one axis of symmetry can be drawn.
Double flowers have an abnormally increased number of petals. In most cases they occur as a result of petals splitting.
Stamen- part of a flower, which is a kind of specialized structure that forms microspores and pollen. It consists of a filament, through which it is attached to the receptacle, and an anther containing pollen. The number of stamens in a flower is a systematic feature. Stamens are distinguished by the method of attachment to the receptacle, by shape, size, structure of the stamen filaments, connective tissue and anther. The collection of stamens in a flower is called the androecium.
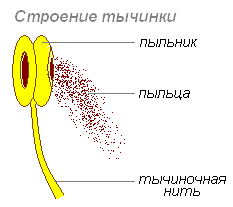
filament- the sterile part of the stamen, bearing an anther at its apex. The filament can be straight, curved, twisted, tortuous, or broken. Shape: hair-like, cone-shaped, cylindrical, flattened, club-shaped. The nature of the surface is bare, pubescent, hairy, with glands. In some plants it is short or does not develop at all.
Anther located at the top of the filament and attached to it by a connective tissue. It consists of two halves connected to each other by a connector. Each half of the anther has two cavities (pollen sacs, chambers, or nests) in which pollen develops.

As a rule, the anther is four-locular, but sometimes the partition between the nests in each half is destroyed, and the anther becomes two-locular. In some plants the anther is even single-locular. Very rarely found with three nests. Based on the type of attachment to the filament, anthers are classified into immobile, movable, and oscillating anthers.
Anthers contain pollen or pollen grains.
Pollen grain structure
The dust particles formed in the anthers of the stamens are small grains; they are called pollen grains. The largest ones reach 0.5 mm in diameter, but usually they are much smaller. Under a microscope you can see that dust particles from different plants are not at all the same. They differ in size and shape.

The surface of the dust particle is covered with various protrusions and tubercles. Once on the stigma of the pistil, the pollen grains are held with the help of outgrowths and the sticky liquid secreted on the stigma.
The nests of young anthers contain special diploid cells. As a result of meiotic division, four haploid spores are formed from each cell, which are called microspores due to their very small size. Here, in the cavity of the pollen sac, microspores turn into pollen grains.

This happens as follows: the microspore nucleus is divided mitotically into two nuclei - vegetative and generative. Areas of cytoplasm are concentrated around the nuclei and two cells are formed - vegetative and generative. On the surface of the cytoplasmic membrane of the microspore, a very strong shell is formed from the contents of the pollen sac, insoluble in acids and alkalis. Thus, each pollen grain consists of vegetative and generative cells and is covered with two membranes. Many pollen grains make up the pollen of a plant. Pollen matures in the anthers at the time the flower opens.
Pollen germination
The beginning of pollen germination is associated with mitotic division, as a result of which a small reproductive cell is formed (sperm cells develop from it) and a large vegetative cell (the pollen tube develops from it).
After the pollen reaches the stigma in one way or another, its germination begins. The sticky and uneven surface of the stigma helps to retain pollen. In addition, the stigma secretes a special substance (enzyme) that acts on pollen, stimulating its germination.
The pollen swells, and the restraining influence of the exine (the outer layer of the pollen grain shell) causes the contents of the pollen cell to rupture one of the pores, through which the intina (the inner, poreless shell of the pollen grain) protrudes outward in the form of a narrow pollen tube. The contents of the pollen cell pass into the pollen tube.
Under the epidermis of the stigma there is loose tissue into which the pollen tube penetrates. It continues to grow, passing either through a special conducting channel between mucus cells, or tortuously along the intercellular spaces of the conductive tissue of the column. In this case, usually a significant number of pollen tubes simultaneously advance in the style, and the “success” of one or another tube depends on the individual growth rate.
Two sperm and one vegetative nucleus pass into the pollen tube. If the formation of sperm cells in pollen has not yet occurred, then a generative cell passes into the pollen tube, and here, through its division, sperm cells are formed. The vegetative nucleus is often located in front, at the growing end of the tube, and sperm are successively located behind it. In the pollen tube, the cytoplasm is in constant motion.

Pollen is rich in nutrients. These substances, especially carbohydrates (sugar, starch, pentosans) are intensively consumed during pollen germination. In addition to carbohydrates in chemical composition Pollen contains proteins, fats, ash and a large group of enzymes. Pollen contains a high phosphorus content. Substances in pollen are in a mobile state. Pollen is easily transferred low temperatures up to - 20Cº and even lower for a long time. High temperatures quickly reduce germination.
Pestle
The pistil is the part of the flower that forms the fruit. It arises from the carpel (a leaf-like structure bearing ovules) subsequently fusion of the edges of the latter. It can be simple if it is made up of one carpel, and complex if it is made up of several simple pistils fused together with side walls. In some plants, the pistils are underdeveloped and are represented only by rudiments. The pistil is divided into ovary, style and stigma.


Ovary- the lower part of the pistil, which contains the seed buds.
Having entered the ovary, the pollen tube grows further and enters the ovule in most cases through the pollen duct (micropyle). Invading the embryo sac, the end of the pollen tube bursts and the contents spill onto one of the synergids, which darkens and quickly collapses. The vegetative nucleus is usually destroyed before the pollen tube penetrates the embryo sac.
Flowers regular and irregular
The tepals (simple and double) can be arranged so that several planes of symmetry can be drawn through it. Such flowers are called regular. Flowers through which one plane of symmetry can be drawn are called irregular.
Flowers bisexual and dioecious
Most plants have flowers that contain both stamens and pistils. These are bisexual flowers. But in some plants, some flowers have only pistils - pistillate flowers, while others have only stamens - staminate flowers. Such flowers are called dioecious.
Monoecious and dioecious plants
Plants that bear both pistillate and staminate flowers are called monoecious. Dioecious plants have staminate flowers on one plant and pistillate flowers on another.
There are species in which bisexual and unisexual flowers can be found on the same plant. These are the so-called polygamous (polygamous) plants.
Inflorescences
Flowers are formed on the shoots. Very rarely they are located alone. Much more often, flowers are collected in noticeable groups called inflorescences. The study of inflorescences began with Linnaeus. But for him, the inflorescence was not a type of branching, but a way of flowering.
Inflorescences are distinguished between main and lateral axes (sessile or on pedicels); such inflorescences are called simple. If the flowers are on the lateral axes, then these are complex inflorescences.
| Inflorescence type | Inflorescence diagram | Peculiarities | Example |
| Simple inflorescences | |||
| Brush | Individual lateral flowers sit on an elongated main axis and at the same time have their own pedicels, approximately equal in length | Bird cherry, lily of the valley, cabbage | |
| Ear | The main axis is more or less elongated, but the flowers are stalkless, i.e. sessile. | Plantain, orchis | |
| cob | It differs from the ear by its thick, fleshy axis. | Corn, calligraphy | |
| Basket | The flowers are always sessile and sit on the strongly thickened and widened end of the shortened axis, which has a concave, flat or convex appearance. In this case, the inflorescence on the outside has a so-called involucre, consisting of one or many successive rows of bract leaves, free or fused. | Chamomile, dandelion, aster, sunflower, cornflower | |
| Head | The main axis is greatly shortened, the lateral flowers are sessile or almost sessile, closely spaced to each other. | Clover, scabiosa | |
| Umbrella | The main axis is shortened; lateral flowers emerge as if from one place, sit on stalks of different lengths, located in the same plane or dome-shaped. | Primrose, onion, cherry | |
| shield | It differs from the raceme in that the lower flowers have long pedicels, so that as a result the flowers are located almost in the same plane. | Pear, spirea | |
| Complex inflorescences | |||
| Complex brush or whisk |  | Lateral branching axes extend from the main axis, on which flowers or simple inflorescences are located. | Lilac, oats |
| Complex umbrella | Simple inflorescences extend from the shortened main axis. | Carrots, parsley | |
| Complex ear | Individual spikelets are located on the main axis. | Rye, wheat, barley, wheatgrass | |
Biological significance of inflorescences
The biological significance of inflorescences is that small, often inconspicuous flowers, when collected together, become noticeable, produce the largest amount of pollen and better attract insects that carry pollen from flower to flower.
Pollination
In order for fertilization to occur, pollen must land on the stigma.
The process of transferring pollen from the stamens to the stigma of the pistil is called pollination. There are two main types of pollination: self-pollination and cross-pollination.
Self-pollination
In self-pollination, pollen from the stamen ends up on the stigma of the same flower. This is how wheat, rice, oats, barley, peas, beans, and cotton are pollinated. Self-pollination in plants most often occurs in a flower that has not yet opened, that is, in a bud; when the flower opens, it is already finished.
During self-pollination, sex cells formed on the same plant and, therefore, having the same hereditary characteristics merge. This is why the offspring produced by the process of self-pollination are very similar to the parent plant.

Cross pollination
During cross-pollination, a recombination of hereditary characteristics of the paternal and maternal organisms occurs, and the resulting offspring can acquire new properties that the parents did not have. Such offspring are more viable. In nature, cross-pollination occurs much more often than self-pollination.

Cross-pollination is carried out with the help of various external factors.
Anemophilia(wind pollination). In anemophilous plants, the flowers are small, often collected in inflorescences, a lot of pollen is produced, it is dry, small, and when the anther opens, it is thrown out with force. Light pollen from these plants can be carried by the wind over distances of up to several hundred kilometers.

The anthers are located on long thin filaments. The stigmas of the pistil are wide or long, feathery and protrude from the flowers. Anemophily is characteristic of almost all grasses and sedges.
Entomophily(transfer of pollen by insects). Adaptations of plants to entomophily are the smell, color and size of flowers, sticky pollen with outgrowths. Most flowers are bisexual, but the maturation of pollen and pistils does not occur simultaneously, or the height of the stigmas is greater or less than the height of the anthers, which serves as protection against self-pollination.
The flowers of insect-pollinated plants have areas that secrete a sweet, aromatic solution. These areas are called nectaries. Nectaries may be in different places flower and have different shapes. Insects, having flown up to a flower, are drawn to the nectaries and anthers and become dirty with pollen during their meal. When an insect moves to another flower, the pollen grains it carries stick to the stigmas.
When pollinated by insects, less pollen is wasted, and therefore the plant conserves nutrients by producing less pollen. Pollen grains do not need to stay in the air for long and can therefore be heavy.
Insects can pollinate sparsely located flowers and flowers in windless places - in the thicket of a forest or in thick grass.
Typically, each plant species is pollinated by several types of insects, and each type of pollinating insect serves several plant species. But there are types of plants whose flowers are pollinated by insects of only one species. In such cases, the mutual correspondence between the lifestyles and structure of flowers and insects is so complete that it seems miraculous.
Ornithophilia(pollination by birds). Characteristic of some tropical plants with brightly colored flowers, abundant nectar secretions, and a strong elastic structure.
Hydrophilia(pollination by water). Observed in aquatic plants. The pollen and stigma of these plants most often have a thread-like shape.
Bestiality(pollination by animals). These plants are characterized by large flower sizes, abundant secretion of nectar containing mucus, and massive production of pollen during pollination. bats- blooms at night.
Fertilization
The pollen grain lands on the stigma of the pistil and is attached to it due to the structural features of the shell, as well as the sticky sugary secretions of the stigma to which the pollen sticks. The pollen grain swells and germinates, turning into a long, very thin pollen tube. The pollen tube is formed as a result of division of a vegetative cell. First, this tube grows between the cells of the stigma, then the style, and finally grows into the cavity of the ovary.
The generative cell of the pollen grain moves into the pollen tube, divides and forms two male gametes (sperm). When the pollen tube penetrates the embryo sac through the pollen duct, one of the sperm fuses with the egg. Fertilization occurs and a zygote is formed.
The second sperm fuses with the nucleus by the large central cell of the embryo sac. Thus, in flowering plants, during fertilization, two fusions occur: the first sperm fuses with the egg, the second with the large central cell. This process was discovered in 1898 by the Russian botanist, academician S.G. Navashin and called it double fertilization. Double fertilization is characteristic only of flowering plants.
The zygote formed by the fusion of gametes is divided into two cells. Each of the resulting cells divides again, etc. As a result of repeated cell divisions, a multicellular embryo of a new plant develops.
The central cell also divides, forming endosperm cells in which reserves accumulate nutrients. They are necessary for the nutrition and development of the embryo. The seed coat develops from the integument of the ovule. After fertilization, a seed develops from the ovule, consisting of a peel, an embryo and a supply of nutrients.
After fertilization, nutrients flow to the ovary, and it gradually turns into a ripe fruit. The pericarp, which protects the seeds from adverse influences, develops from the walls of the ovary. In some plants, other parts of the flower also take part in the formation of the fruit.

Education dispute
Simultaneously with the formation of pollen in the stamens, the formation of a large diploid cell occurs in the ovule. This cell divides meiotically and gives rise to four haploid spores, which are called macrospores because they are larger in size than microspores.

Of the four macrospores formed, three die, and the fourth begins to grow and gradually turns into an embryo sac.
Formation of the embryo sac
As a result of threefold mitotic division of the nucleus, eight nuclei are formed in the cavity of the embryo sac, which are covered with cytoplasm. Cells deprived of membranes are formed, which are arranged in a certain order. At one pole of the embryo sac, an egg apparatus is formed, consisting of an egg and two auxiliary cells. At the opposite pole there are three cells (antipodes). One nucleus migrates from each pole to the center of the embryo sac (polar nuclei). Sometimes the polar nuclei fuse to form the diploid central nucleus of the embryo sac. The embryo sac in which nuclear differentiation has occurred is considered mature and can receive sperm.

By the time the pollen and embryo sac mature, the flower opens.
Structure of the ovule
Ovules develop on internal sides the walls of the ovary and, like all parts of the plant, consist of cells. The number of ovules in the ovaries of different plants varies. In wheat, barley, rye, and cherry, the ovary contains only one ovule, in cotton - several dozen, and in poppy, their number reaches several thousand.
Each ovule is covered with a cover. At the top of the ovule there is a narrow canal - the pollen passage. It leads to the tissue that occupies the central part of the ovule. In this tissue, as a result of cell division, an embryo sac is formed. Opposite the pollen opening there is an egg cell, and the central part is occupied by a large central cell.
Development of angiosperms (flowering) plants

Formation of seed and fruit
When the seed and fruit are formed, one of the sperm fuses with the egg, forming a diploid zygote. Subsequently, the zygote divides many times, and as a result, a multicellular plant embryo develops. The central cell, fused with the second sperm, also divides many times, but the second embryo does not arise. A special tissue is formed - endosperm. The endosperm cells accumulate reserves of nutrients necessary for the development of the embryo. The integument of the ovule grows and turns into a seed coat.

Thus, as a result of double fertilization, a seed is formed, which consists of an embryo, storage tissue (endosperm) and a seed coat. The wall of the ovary forms the wall of the fruit, called the pericarp.
Sexual reproduction
Sexual reproduction in angiosperms is associated with flowers. Its most important parts are the stamens and pistils. Complex processes associated with sexual reproduction occur in them.
In flowering plants, male gametes (sperm) are very small, while female gametes (eggs) are much larger.
In the anthers of the stamens, cell division occurs, resulting in the formation of pollen grains. Each pollen grain of angiosperms consists of vegetative and generative cells. The pollen grain is covered with two coats. The outer shell, as a rule, is uneven, with spines, warts, and mesh-like outgrowths. This helps the pollen grains to stay on the stigma. The pollen of a plant, ripening in the anthers, consists of many pollen grains by the time the flower blooms.
Flower formula
Formulas are used to conditionally express the structure of flowers. To compile a flower formula, use the following notation:
A simple perianth consisting of only sepals or only petals; its parts are called tepals. |
|
| H | Calyx, consists of sepals |
| L | Corolla, consists of petals |
| T | Stamen |
| P | Pestle |
| 1,2,3... | The number of flower elements is indicated by numbers |
| , | Identical parts of a flower, differing in shape |
| () | Fused parts of a flower |
| + | Arrangement of elements in two circles |
| _ | Upper or lower ovary - a line above or below the number that shows the number of pistils |
| Wrong flower | |
| * | The right flower |
| ♂ | Unisexual staminate flower |
| ♀ | Unisexual pistillate flower |
| Bisexual | |
| ∞ | Number of flower parts exceeding 12 |
An example of a cherry blossom formula:
*H 5 L 5 T ∞ P 1
Flower diagram
The structure of a flower can be expressed not only by a formula, but also by a diagram - a schematic representation of a flower on a plane perpendicular to the axis of the flower.

Make a diagram using cross sections of unopened flower buds. The diagram gives a more complete idea of the structure of a flower than a formula, since it also shows the relative position of its parts, which cannot be shown in a formula.
Pollen grain pollen grain- pollen grain.
The male gametophyte of a seed plant, developing from a microspore and limited by its shell; shell P.z.(like microspores) consists of 2 layers - the outer (exine) and the inner (intine) very diverse in structure; dimensions P.z. from 2 to 250 microns.
(Source: “English-Russian explanatory dictionary of genetic terms.” Arefiev V.A., Lisovenko L.A., Moscow: VNIRO Publishing House, 1995)
See what a “pollen grain” is in other dictionaries:
- (speck of dust) male gametophyte of seed plants, in which, at the time of pollination or after it, male reproductive sperm cells (or spermatozoa) necessary for fertilization develop. Develops from microspores into microsporangia. The totality... ... Big Encyclopedic Dictionary
Speck of dust (granum pollinis), male. gametophyte of a seed plant. Development begins from a microspore in a microsporangium and completes it after pollination, i.e. transfer to the pollen chamber of the ovule (in gymnosperms) or on the stigma of the pistil (in ... ... Biological encyclopedic dictionary
pollen grain- Male gametophyte of a seed plant, developing from a microspore and limited by its shell; shell P.z. (like microspores) consists of 2 layers, very diverse in structure, outer (exine) and inner (intine); dimensions of P.Z. from 2 to... ... Technical Translator's Guide
Pollen grain- * pollen grain * pollen grain male gametophyte (q.v.) of a seed plant, developing from microspores (q.v.), which in turn are formed from the microsporocyte, or pollen mother cell, during two divisions of meiosis (q.v.) and contain, in addition... ... Genetics. encyclopedic Dictionary
- (speck of dust), the male gametophyte of seed plants, in which, at the time of pollination or after it, male reproductive sperm cells (or spermatozoa) necessary for fertilization develop. Develops from microspores into microsporangia. The totality... ... encyclopedic Dictionary
pollen grain- PLANT EMBRYOLOGY POLLEN grain is a male gametophyte, consisting of vegetative and generative cells and two membranes (intine and exine). In many plant species, the generative cell in the pollen grain divides, forming two sperm cells... General embryology: Terminological dictionary
pollen grain- žiedadulkė statusas T sritis augalininkystė apibrėžtis Sėklinių augalų vyriškasis gametofitas. atitikmenys: engl. pollen grain rus. pollen grain... Žemės ūkio augalų selekcijos ir sėklininkystės terminų žodynas
Dust mote, male gametophyte of a seed plant; begins development from a microspore (See Microspore) in microsporangium and completes it after pollination, i.e., transfer of the ovule to the pollen chamber (in gymnosperms) or on the stigma of the pistil (in ... ... Great Soviet Encyclopedia
- (speck of dust), husband. gametophyte of seed plants, in which the males develop at the time of pollination or after it. germ cells sperm (or sperm) necessary for fertilization. Develops from microspores into microsporangia. The totality of P. z. pollen... Natural science. encyclopedic Dictionary
pollen grain- Synonyms: speck of dust, male gametophyte of seed plants; develops from microspores into microsporangia (see Fig. Pollen grains: a - pea, b - cornflower, c - rapeseed) ... Anatomy and morphology of plants
Functionally, a pollen grain is a biological container that contains the male gametophyte of angiosperms and gymnosperms. Such a container protects male gametes from adverse environmental influences during their transfer from the anthers to the pistil.
Pollen grains various types plants vary both in size (from 10 to almost 100 microns) and in shape: round, oval, disc-shaped, bean-shaped, and also filamentous. The natural color of pollen is mostly white, cream, yellow or orange. The texture of the cell wall of pollen also varies, from smooth to covered with various outgrowths.
The pollen grain shell consists of two main layers - exine and intine (Fig. 2). The structure of these two layers, as well as their thickness, are important taxonomic characteristics of plants.
Intina is the inner layer of the pollen grain shell. It encloses its contents and serves as the material that forms the pollen tube. Its chemical composition is heterogeneous: the outer layer is formed mainly by pectin, while the inner layer consists mainly of cellulose and pectin. Intine is easily destroyed by acids and alkalis and is not preserved in a fossil state. Intina is present in pollen grains of all flowering plants.
Exine is the outer layer of the pollen grain coat. Thanks to its main component, sporopollenin, exine is characterized by extraordinary durability: unlike intine, it does not dissolve in acids and alkalis, can withstand temperatures up to 100°C and is preserved for millions of years in geological sediments. Sporopollenin protects the vegetative and generative cells of pollen grains from mechanical damage, chemical breakdown, drying and ultraviolet radiation. On the surface of the outer wall you can sometimes find grains covered with a liquid, fatty substance, the so-called “pollenkit”.
Exine consists of two main layers - ectexine and endexine, to which in many plants is added a middle layer - mesexine.
Ecticine (ectexina, from the Greek ektos - outside, outside), or external exine, is the most stable part of the exine and is distinguished by its exceptional diversity of structure. In flowering plants, there are 3 main types of ectexine (Fig. 3): columnar or columellate (also called baculate), granular and homogeneous.
Figure 2. Pollen grain structure. 1 – ectexine, 2 – intine, 3 – endexine, 4 – nucleus of a vegetative cell, 5 – vegetative cell, 6 – aperture, 7 – generative cell, 8 – nucleus of a generative cell
Columnar ectexine is very widespread in flowering plants and is found almost exclusively in them. Usually the columns are covered on top with a roof-like cover, which in palynology is designated special term“tectum” (from Latin tectum - roof, roof, canopy), or cover, less often ectexine coverless (intectate). The columns are of many types and often reach great complexity, branching repeatedly at the top. They rise on a continuous underlying layer called the substratum.
Granular ectexine is much less typical for flowering plants. It occurs with or more often without a tectum, with or without an underlying layer.
Homogeneous ectexine is very rare, characteristic of some primitive genera, such as degeneration, etc. Along with the structure of ectexine, i.e. internal structure, the nature of the sculpture of its surface is also very important. There are numerous types of surface sculptures. Many sculptural elements are so small (less than 0.1 microns) that they can only be detected using an electron microscope (Fig. 4). It should be noted that the allergenic properties of the pollen grain shell have been practically not studied.
Along with the structure of ectexine, i.e. its internal structure, the nature of the sculpture of its surface is also important. There are numerous types of surface sculptures. Many sculptural elements are so small (less than 0.1 microns) that they can only be detected using an electron microscope (Fig. 4).
Endexina (endexina, from the Greek endon - inside), or internal exine, is usually much thinner than ectexine, often very thin and often even absent. Like ectexine, it is resistant to acids and alkalis, but unlike it, it is not stained with basic fuchsin. While ectexine consists mainly of radial structural elements, the structure of endexine is lamellar (lamella), granular (granular) or often homogeneous.
As a rule, in the exine of a pollen grain there are elastic, flexible, often thin or even perforated places that serve for the exit of the pollen tube - the aperture. The structure of apertures, their number and location on the surface of the pollen grain are the most important features in establishing the morphological types of pollen.
Pollen is often collected in dyads (Scheuchzeria), tetrads (cattails), polliads (Mimosa), pollinia (Orchids) (Fig. 5).
The strengthening of tetrads or the cohesion of individual pollen grains is also facilitated by viscine filaments attached to or near the proximal pole of the grains. The length of the thread reaches 1500 microns. Viscine threads are characteristic of the pollen of several families of insect-pollinated plants: heathers, fireweeds, proteaceae, and some nymphaeaceae. The structure of pollen grains undoubtedly has great importance for the manifestation or absence of allergenic properties of pollen different types plants, however, very little has been studied in this aspect to date.
The search for new allergens remains relevant; in particular, work is actively underway to study the allergenic composition of the surface of pollen grains. It is believed that the surface components of pollen contribute to the development of allergenic diseases, but in general, the allergenic properties of the pollen surface have been poorly studied. This is mainly due to the fact that upon receipt, pollen extracts undergo a stage of washing with organic solvents in order to get rid of possible contamination with pollutants and microorganisms. As a result, the surface molecules of the inner cellulose layer (intin), outer wall (exine), and extracellular matrix containing lipophilic proteins and lipids are washed off the pollen grains. Therefore, when identifying pollen allergens using soluble protein extracts, the surface components of pollen grains remain undetected.
Vigh-Conrad K.A. et al. analyzed the level of sIgE in various sera in a microarray format and compared the allergenic properties of various pollen fractions (surface, cytoplasmic fractions and commercial pollen preparations). The microarrays contained surface and commercial pollen extracts of 22 plant species, commercial extracts of 9 non-pollen allergens, and 5 recombinant allergenic proteins. To separate the surface and cytoplasmic fractions, samples were extracted with organic solvents or aqueous buffers, respectively. Allergens in microchips were incubated with the test serum, after which the level of sIgE was assessed by indirect immunofluorescence. Screening demonstrated that such microarray analysis well reflects serum sIgE levels. A difference was found in the allergenic composition of surface and cytoplasmic pollen fractions.
From a microspore and performing the function of pollination, that is, fertilization of the female gametophyte located in the ovule.
Structure
Pollen develops in the anthers of the stamens. The layer of the young anther bordering the outer skin is divided by partitions into two layers, of which the outer one produces the wall of the pollen chamber, and the inner one gives rise to archesporium, consisting of producing, or mother, pollen cells. Then the pollen mother cells usually increase in size and thicken the shell. They either remain connected to each other (most dicotyledons) or separate (many monocotyledons). Each producing cell then divides into four specifically producing cells, either through repeated division into two (in monocots), or around those resulting by dividing four nuclei, the shells of four cells appear at once (in most dicotyledons). The contents inside each special producing cell are clothed with a new shell, the sporoderm, which differentiates into an outer thick layer, exine, and an inner layer, intina. The spores of horsetails, ferns and mosses are characterized by another layer above the exine, called the featherbed or perisporium. The science that studies sporoderm is called palynology, while the inner part of pollen is studied by embryology.
For the most part, the resulting pollen cells are soon completely separated, sometimes they remain connected in fours (tetrads, or quadruples), for example, in many orchids (cache, nest), cattail, annona, rhododendron, etc. In orchids from the tribe Orchideae, pollen cells are connected in large numbers into pollen bodies (lat. massulae), which, in turn, are interconnected into one mass, the so-called pollinarium, filling the entire pollen chamber. The same is observed in many Asclepiaceae.
The size of pollen cells ranges from 0.0025 to 0.25 mm. They are predominantly ellipsoidal or spherical, sometimes faceted or angular. The outer layer, exine, is often covered with various sculptures in the form of ridges, tubercles, spines, etc., sometimes dry and smooth. In the sporoderm there may be through hole or an easily extensible area called the aperture. The pollen of many conifers has air sacs, which are outgrowths of the sporoderm and serve to facilitate the flight of pollen.
Pollen as a transmitter of environmental information in plants
According to the ideas existing in classical genetics, pollen carries only genetic information, and the amount of pollen falling on female flower, does not play a role, since one pollen grain is enough for fertilization. In 1977, V. Geodakyan suggested that the amount of pollen falling on a female flower can also carry environmental information about the situation in the habitat. A large number of pollen indicates optimal environmental conditions (area center, a lot male plants, good conditions for growth and weather), while a small amount of pollen, on the contrary, carries information about unfavorable conditions: this happens either on the periphery, where population density drops sharply, or in the center, when extreme conditions occur there, which eliminate primarily male individuals.
Pollen quantity can determine the sex ratio, dispersion, and sexual dimorphism of a plant population. A large amount of pollen leads to a decrease in these characteristics and stabilization of the population. A small number leads to their increase and destabilization of the population.
The dependence of the secondary sex ratio on the amount of pollen was confirmed in four species of dioecious plants belonging to three families - Sour sorrel (Buckwheat), White slumber (Cloveaceae), Hemp and Japanese hop (Hemp). As the amount of pollen increases, the number of male plants in the offspring decreases and vice versa. The amount of pollen depends on the density of planting, so in dense plantings the proportion of female plants should increase.
The role of pollen in the life of bees
After passing through the bees' feet, the pollen becomes pollen, that is, pollen that the bees have collected and processed with their enzymes.
Thanks to pollen, the mass of yesterday's larva increases hundreds of times in just a few days, the wings become stronger and straighten, and all the working glands are formed. Pollen also serves as raw material for the creation of royal jelly, a product intended to feed the queen.
In a bee colony, pollen is primarily needed by nurse bees. They intensively eat this protein-lipid food, which is necessary for the production of royal jelly, which feeds the young queen bee and, in the first 3 days, the larvae of worker bees. Newly born bees also feed on pollen: their bodies have little nitrogen, they need proteins and vitamins. Builder bees need pollen for the functioning of their wax glands, and drones need it for normal puberty and functioning. During the season, a bee colony collects and consumes 35-40 kg of pollen.
Microbiota of pollen grains
It has been shown that pollen grains of different plant species are characterized by a specific set of bacteria living on their surface (microbiome).
Organization of the microbiota of pollen grains
It is significant that the forms cultivated in laboratory conditions constitute a minority of the entire microbiota of pollen grains (for the above plants - 44 species, that is, less than 5%). However, there are quite a lot of cultivated bacteria on the surface of pollen grains - from 10 6 (silver birch) to 10 9 (autumn colchicum) cells per gram of pollen.
Studying the surface of pollen grains using scanning electron microscopy makes it possible to identify bacterial colonies, including those organized in the form of biofilms.
Interaction of plants with their own microbiota
It has been shown that the plant interacts with its own microbiome, synthesizing, in particular, antimicrobial peptides. a nti m icrobial p eptides, AMPs) . Among plant antimicrobial peptides, defensin-like peptides are the most studied and, apparently, the most widespread. Thus, in the rhizoid moth Tal ( Arabidopsis thaliana) of approximately 800 cysteine-rich peptide genes, about three hundred genes are defensin-like peptide genes. Defensins are characterized by the presence of a specific tertiary structure with a disulfide bridge between cysteine residues.
The importance of antimicrobial peptides in pollen is indicated by the fact that 80 of the 300 defensin-like peptides Arabidopsis thaliana homologous to the defensin-like peptide PCP-A1, previously isolated from the pollen shell of cabbage ( Brassica) . It has been shown that a number of antimicrobial peptides prevent self-pollination by interacting with receptors in pistil tissues, but their effect on the microbiota of pollen grains has been studied very little.
U Arabidopsis thaliana at least 200 of the 800 cysteine-rich peptide genes have been shown to be expressed in the embryo sac. A hypothesis has been proposed that these peptides provide an antimicrobial barrier for the embryo sac. Perhaps this barrier isolates the embryo sac from bacteria that can penetrate it during the growth of the pollen tube through the ovary.
It should be clarified that functions of some antimicrobial peptides that are not related to protection against microorganisms and self-recognition during pollination have also been identified. A number of peptides that are produced in the synergid cell and direct the growth of the pollen tube have been studied; antimicrobial peptides that initiate rupture of the pollen tube upon entering the embryo sac have also been identified.
The role of the pollen microbiome in interactions with animals
The interaction of the pollen microbiome with pollinators, as well as with the human immune system during an allergic reaction to pollen, remains poorly understood.
Literature
- Sladkov A. N. Morphology of pollen and spores of modern plants in the USSR in connection with methods of its practical application. - M.: Moscow University Publishing House, 1962. - P. 256.
- Sladkov A. N. Introduction to spore-pollen analysis. - M.: Nauka, 1967.
- Kosenko Ya. Microworld of pollen // Floriculture. - 2008. - No. 4. - pp. 16-17.
see also
Notes
- Pollen // Great Soviet Encyclopedia: [in 30 volumes] / ch. ed. A. M. Prokhorov. - 3rd ed. - M.: Soviet Encyclopedia, 1969-1978. (Retrieved December 8, 2013)
- Meyer N.R. Sporoderm // Plant life: in 6 volumes / ch. ed. Al. A. Fedorov. - M.: Education, 1978. - T. 4: Mosses. Moss mosses. Horsetails. Ferns. Gymnosperms / ed. I. V. Grushvitsky and S. G. Zhilin. - pp. 29-32. - 447 p. - 300,000 copies.
- Geodakyan V. A. (1977) Pollen quantity as a regulator of the evolutionary plasticity of cross-pollinating plants. DAN USSR, vol. 234 , No. 6. p. 1460-1463.
- Geodakyan V. A. (1978) Pollen quantity as a transmitter of environmental information and a regulator of plant evolutionary plasticity. Zhypn. total biology. 39 No. 5, p. 743-753.
- Correns S. (1922) Geschlechtsbestimmung und Zahlenverhaltnis der Geschlechter beim Sauerampfer (Rumex acetosa). "Biol. Zbl." 42 , 465-480.
- Rychlewski J., Kazlmierez Z. (1975) Sex ratio in seeds of Rumex acetosa L. as a result of sparse or abundant pollination. "Acta Biol. Cracov" Scr. Bot., 18 , 101-114.
- Correns C. (1928) Bestimmung, Vererbung und Verteilung des Geschlechter bei den hoheren Pflanzen. Handb. Vererbungswiss., 2 , 1-138.
- Mulcahy D. L. (1967) Optimal sex ratio in Silene alba. "Heredity" 22 № 3, 41.
- Riede W. (1925) Beitrage zum Geschlechts- und Anpassungs-problem. "Flora" 18/19
- Kihara H., Hirayoshi J. (1932) Die Geschlechtschromosomen von Humulus japonicus. Sieb. et. Zuce. In: 8th Congr. Jap. Ass. Adv. Sci., p. 363-367 (cit.: Plant Breeding Abstr., 1934, 5 , No. 3, p. 248, ref. No. 768).
- Geodakyan V. A., Geodakyan S. V. (1958) Is there a negative Feedback in determining gender? Journal of General Biology. 46 No. 2, p. 201-216.
- Ter-Avanesyan D. V. (1949) Tr. according to adj. botany, genetics and selection. 28 119.
- Ter-Avanesian D. V. (1978) Significance of pollen amount for fertilization. "Bull.Torrey Bot.Club." 105 N 1, 2-8.
- Khismatullina N.Z. Apitherapy. - Perm: Mobile, 2005. - P. 74.
- H. Colldahl, L. Nilsson. Possible relationship between some allergens (pollens, mites) and certain microorganisms (bacteria and fungi). A morphological study using the scanning electron-microscope // Acta Allergologica. - October 1973. - T. 28, issue. 4 . - pp. 283–295. - ISSN 0001-5148.
- Robert R. Junker, Alexander Keller. Microhabitat heterogeneity across leaves and flower organs promotes bacterial diversity // FEMS Microbiology Ecology. - 2015-09-01. - T. 91, issue. 9 . - ISSN 0168-6496. - DOI:10.1093/femsec/fiv097.
- Andrea Obersteiner, Stefanie Gilles, Ulrike Frank, Isabelle Beck, Franziska Häring. Pollen-Associated Microbiome Correlates with Pollution Parameters and the Allergenicity of Pollen // PLOS ONE. - 2016-02-24. - T. 11, issue. 2. - S. e0149545. - ISSN 1932-6203. - DOI:10.1371/journal.pone.0149545.
pollen grain
POLLEN GRAIN (speck of dust) is a male gametophyte of seed plants, in which, at the time of pollination or after it, male reproductive cells develop - sperm (or spermatozoa), necessary for fertilization. Develops from microspores into microsporangia. The collection of pollen grains is pollen.
Pollen grain
mote, male gametophyte of a seed plant; begins development from a microspore in a microsporangium and completes it after pollination, i.e., transfer of the ovule to the pollen chamber (in gymnosperms) or on the stigma of the pistil (in angiosperms). P. z. has 2 shells: outer ≈ exine, strong and resistant, and inner ≈ intine, consisting mainly of fiber and pectin substances. The exine usually has thin areas or openings, the so-called. apertures are grooves or pores through which, during the germination of P. z. the protoplast covered with intine protrudes, i.e., pollen tubes form.
In gymnosperms P. z. by the time of pollination, it consists of several living cells (in some, also from the remains of dead ones), among which there are vegetative (haustorial) and generative (antheridial); the first of them in the pollen chamber forms a pollen tube (haustorium), which penetrates the nucellus, the second, dividing, forms a spermatogenic, or spermiogenic, cell and a sister cell (the so-called stalk). The spermatogenic cell then forms gametes - multiflagellate sperm (for example, in cycads) or flagellated sperm (for example, in conifers), which reach the archegonia of the female prothallus through pollen tubes. In angiosperms, by the time P. z. reaches the stigma of the pistil. consists of either a siphonogenic cell and a spermiogenic cell located inside it (bicellular or binucleate pollen), or, if the spermiogenic cell has already divided, ≈ 2 sperm cells located inside the siphonogenic cell (tricellular or trinuclear pollen). During the germination of three-celled P. z. the nucleus of the siphonogenic cell and both sperm enter the pollen tube; during bicellular germination ≈ the nucleus of a siphonogenic cell and a spermiogenic cell, which divides into 2 sperm in the pollen tube. Development of P. z. ends when the pollen tube reaches the embryo sac, into which both sperm participating in double fertilization enter.
In most seed plants P. z. single (monads); in some angiosperms there are microspores and P. z. developing from them. connected by 2 (dyads; for example, in Scheuchzeria), by 4 (tetrads; in many heathers, some orchids, etc.), by 8≈12≈16≈32 (polyads; in Mimosa); in the swallowtails and some orchids, all the parasites remain connected. one or two anther nests (so-called pollinia).
The shape, size, and structure of the aperture, especially the structure of its exine, the sculpture of the surface, the structure of the apertures and their position are varied, but constant in plants of the same species; and among representatives of different taxa, as a rule, the more similar they are, the closer their relationship. Therefore, the study of P. z. important for plant taxonomy (see Palynology). Due to the persistence of exine, which is well preserved in sedimentary rocks, study of P. z. is the basis of one of the methods of paleobotanical research - spore-pollen analysis.